


Стабильность и аутолиз нейронов коры головного мозга взрослых крыс после смерти
Сергей В. Шелег 1, Джонин Р. ЛоБелло 2, Хью Хиксон 1, Стефен У. Кунс 2, Дэвид ЛауриDavid 3, Михаил К. Недзвед 4
Аффиляция: 1. Фонд продления жизни Alcor, Скоттсдейл (Аризона, США); 2. Отделение патологий, госпиталь и медицинский центр св. Иосифа, Феникс (Аризона, США); 3. факультет биологии университета штата Аризона, Темпе (Аризона США) и 4. отделение патологий Минского государственного медицинского института, Минск (республика Беларусь). Получено 31 июля 2007 года; одобрено и опубликовано в сети Интернет 30 августа 2007 года.
Абстракт
Мы изучали динамику аутолитических повреждений нейронов коры головного мозга, появляющихся у взрослых крыс в течение 24 часов при комнатной температуре (+20ºC) после остановки сердца. Прогрессивные гистологические и ультраструктурные изменения были выявлены при помощи традиционного и имуногистохимического окрашиваний с последующим изучением препаратов под электронным микроскопом. Полученные нами результаты свидетельствуют о том, что в первые шесть часов при комнатной температуре после остановки сердца аутолитических повреждений в ультраструктуре нейронов мозга не наблюдается, что согласуется с предыдущими исследованиями, проведенными на других млекопитающих. Интересно, что после 9 часов после остановки сердца в значительном количестве нейронов мозжечка и неокортекса наблюдалась активизация каспазы-3. При окрашивании и аминокислотой с медью, и по методу Ниссля (с использованием тионина) не было замечено никаких значительных изменений.
Ключевые слова: смерть мозга, аутолиз, церебральная гипоксия, апоптоз, каспаза-3.
Введение
Смерть - это прекращение всех жизненных (обменных) процессов. Соматическая смерть характеризуется прекращением сердечной и дыхательной деятельности, что приводит к гибели всех клеток тела из-за дефицита кислорода. Во многих странах, в том числе в США, смерть мозга приравнивается к юридической смерти [1].
Существует лишь несколько научных публикаций, посвященных ультраструктурным изменениям нейронов центральной нервной системы во время длительной гипоксии головного мозга [2, 3, 4, 5]. К сожалению, в этих опубликованных исследованиях не затрагивалась проблема динамики аутолитических повреждений нейронов ЦНС. Целью нашего исследования является изучение прогрессивных изменений нейронов коры мозга при длительной аноксии всего головного мозга, вызванной остановкой сердца при комнатной температуре. В оцениваемые параметры вошли морфологические (микроскопические и ультраструктурные) и биохимические изменения. Хотя некоторые из этих параметров уже были ранее исследованы на нескольких животных-моделях, но все они проводились либо на мозге новорожденных животных [6], либо в условиях гипотерми [7, 8], либо с проведением реперфузии до вскрытия [9]. Наше исследование, возможно, является первым исследованием мозга взрослых грызунов при нормальной комнатной температуре по нескольким параметрам.
Материалы и методы
Животные
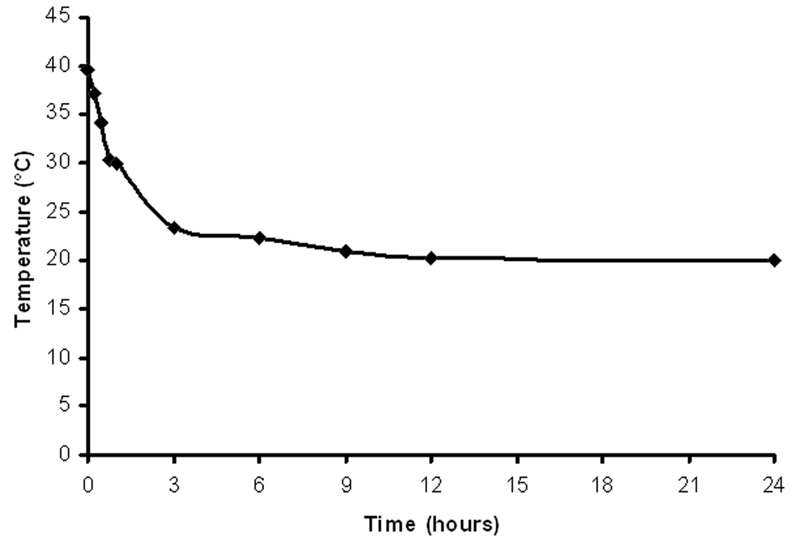
Животные (18 крыс Wistar, самцы, возраст: 94-112 дней, вес: 375-399 г.) были закуплены у Harlan Inc. (Индиаполис, штат Индиана). Животные были усыплены с помощью галотана (Sigma-Aldrich; Cat. #B4388), затем их тела хранились при комнатной температуре (КТ) (+20°C). Мозг животных препарировался по прошествии 1, 3, 6, 12 и 24 часов после остановки сердца при комнатной температуре. Протокол исследования был одобрен Институциональным комитетом Алькора по уходу за животными и их использованию.
Электронная микроскопия
Образцы коры головного мозга (3×3×3 мм) были взяты с лобной доли мозга и зафиксированы 2.5% глутаральдегидом в 0.1M натриевом буфере Соренсена (Electron Microscopy Sciences; Cat. #15980). Затем была выполнена пост-фиксация образцов 1% оксидом осмия в том же самом буфере в течении 1 часа при комнатной температуре, после они были промыты 3 раза в том же буфере и 3 раза в дистиллированной воде. Совместное окрашивание образцов 0.5% уранилацетатом проводилось в течении 2 часов при комнатной температуре, затем образцы были промыты 3 раза дистиллированной водой и, перед помещением в ацетон, были постепенно дегидрированы этанолом. Для инфильтрации использовалась эпоксидная смола Спурра. Затем образцы полимеризировались в течении 48 часов при температуре 60°C. С помощью микротома Leica Ultracut R были нарезаны тонкие срезы, которые были ещё раз окрашены уранилацетатом и цитратом свинца. Образцы были изучены и сфотографированы при ускоряющемся напряжении 80 кВ с помощью Philips CM-12 Scanning Transmission Electron Microscope (Голландия) в лаборатории электронной микроскопии и биовизуализации У. М. Кека Университета штата Аризона.
Световая микроскопия
Мозг животных был зафиксирован в 10% фосфатном буферном формалине (Fisher; Cat. #SF100–4). После мозг был обработан 20% глицерином и 2% диметилсульфоксидом для предотвращения повреждений при замораживании и введен в желатиновый матрикс при помощи технологии MultiBrainTM (NeuroScience Associates, Ноксвил, Теннеси). После отвердевания блок был быстро заморожен погружением в изопентан, предварительно охлаждённый до –70°C дроблёным сухим льдом, и помещён на предметный столик для замораживания нарезающего микротома AO 860. Блок MultiBrainTM был фронтально разделен на части в 35 мкм. Образцы мозга были полностью нарезаны и последовательно помещены в сетку контейнеров 4×5, часть из которых затем была заполнена 10% фосфатным буферным формальдегидом, а часть раствором для сохранения антигенов (натрийфосфатный буфер pH 7.0 - 50%, этиленгликоль - 50%, поливинилпирролидон - 1%) для последующего окрашивания гематоксилином и эозином, специального и иммуногистохимического окрашивания. Для выявления каспазы-3 были проведены окрашивания гематоксилином и эозином, серебром, нейтральным красным, тионином (по методу Ниссля) и иммунологическое окрашивание около 176 отдельных срезов, сделанные через интервалы, равные 1920 мкм [10]. Препараты были изучены и сфотографированы в лаборатории биовизуализации У. М. Керка биологического факультета Университета штата Аризона.
Иммуногистохимия каспазы-3
Для иммуногистохимии каспазы-3 были окрашены свободноплавающие срезы. После обработки пероксидом водорода и блокирующей сывороткой, срезы были подвергнуты иммунологическому окрашиванию при помощи раствора из (1:600) первичного поликлонального антитела на каспазу-3 (Pharmingen, каталог #557035), вторичного антитела и авидин-биотин-HRP (пероксидаза хрена) комплекса (Vectastain ABC Elite Kit, каталог #PK-6101). Инкубационные периоды составили: 1 час для блокирующего антитела, 24 часа для первичного антитела, 1 час для вторичного антитела и 2 часа для авидин-биотин-HRP комплекса. Все инкубации были проведены при комнатной температуре. Срезы были обработаны тетрагидрохлорид диаминобензидином, окисленным пероксидом водорода для визуализации сайтов связывания антител и закреплены на предметных стеклах, покрытых желатином в качестве подложки.
Результаты
Температура животных была измерена с помощью глубоко введенного пищевого зонда. График, показывающий температуру тел животных в разные периоды времени после остановки сердца (Рис. 1).
Рис. 1
Электронная микроскопия
Данные электронной микроскопии не показали очевидных аутолитических изменений нейронов коры в первые 6 часов после остановки сердца при комнатной температуое (Рис. 2А). Спустя 3 часа после общей нормотермической аноксии головного мозга, было обнаружено набухание митохондрий, причиной которого, возможно, стало изменение проницаемости мембран (Рис. 2В). Вакуолизация грубого эндоплазматического ретикулума (RER) и набухание лизосом наблюдались только спустя 6 часов после остановки сердца. Спустя 6 часов после аноксии головного мозга при комнатной температуре ультраструктура митохондриального гребешка не была повреждена. В митохондриях не было обнаружено никаких признаков уменьшения объема, увеличения плотности матрикса и отложения электронно-плотного материала. В митохондриях перинуклеарных зон некоторых нейронов были зафиксированы небольшой отек матрикса и внутреннее набухание гребешка. Спустя 6 часов в нейронах было зафиксировано скопление хроматина. (Рис. 2С). Первый признак аутолиза - исчезновение рибосом - наблюдался только спустя 9 часов при комнатной температуре после остановки сердца всего лишь у 55% нейронов (рис. 2D). Спустя 24 часа после аноксии мозга при комнатной температуре в около 72% нейронов коры головного мозга крыс наблюдался полный хроматолиз (Рис. 2F).
Рис. 2

На рисунке 2:
- А. Состояние нейронов коры мозга крысы спустя час после остановки сердца при комнатной температуре. Неповрежденные клеточные ультраструктуры (масштаб — 0.5мкм).
- B. Состояние нейронов коры мозга крысы спустя 3 часа после остановки сердца при комнатной температуре. В целом митохондрии не повреждены, только что появились некоторые признаки их набухания (черные стрелки) (масштаб — 0.5 мкм).
- С. Нейроны коры головного мозга крысы через 6 часов после остановки сердца при комнатной температуре с вакуолизацией грубого эндоплазматического ретикулума (черные стрелки) (масштаб — 0.5 мкм).
- D. Состояние нейронов коры мозга крысы спустя 9 часов после остановки сердца при комнатной температуре. Митохондрии и ядерная оболочка нейрона не повреждены. Заметны вакуолизация грубого эндоплазматического ретикулума, исчезновение рибосом, первичные признаки аутолиза и набухание лизосом (черные стрелки) (масштаб — 1.0 мкм).
- E. Состояние нейронов коры мозга крысы спустя 12 часов после остановки сердца при комнатной температуре. Присутствует аутолиз и набухание лизосом (черные стрелки) (масштаб — 0.5 мкм).
- F. Состояние нейронов коры мозга крысы спустя 24 часа после остановки сердца при комнатной температуре. Заметны клеточный аутолиз и хроматолиз (черные стрелки) (масштаб — 1.0 мкм).
Световая микроскопия
Обычное окрашивание гематоксилином и эозином не выявило существенных гистологических изменений нейронов коры, мозжечка и гиппокампа в течение периода исследования (Рис. 3A и 3B).
Рис. 3
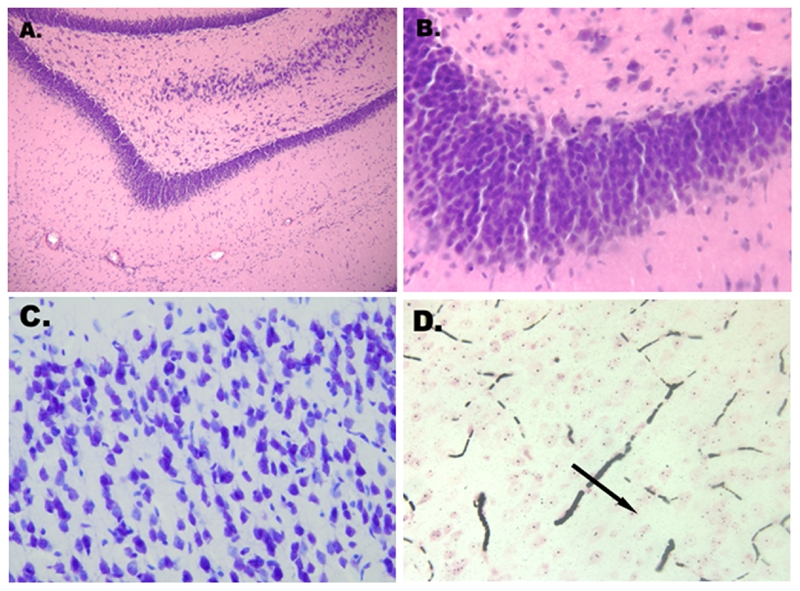
Рис. 3А и В. Отсутствие существенных гистологических изменений нейронов гиппокампа спустя 12 часов после остановки сердца при комнатной температуре (гематоксилин - эозин ×100; ×400). С. Отсутсвие пиктонических нейронов спустя 9 часов после остановки сердца при комнатной температуре (Ниссль,×400). D. Монетные столбики (черная стрелка) эритроцитов (густая кровь) в микроциркуляции в коре головного мозга спустя 1 час после остановки сердца при комнатной температуре (окрашивание серебром,×400).
Окрашивание тионином (по методу Ниссля) не выявило никаких ядерных повреждений, целостность ядра и вещество Ниссля не нарушились в течении всего времени исследования. Окрашивание по методу Ниссля выявило хроматолиз в клетках Пуркинье и пирамидных клетках гиппокампа спустя 12 часов после остановки сердца при комнатной температуре. Спустя 9 часов после аноксии мозга при комнатной температуре гиперхроматоза или пикнотических нейронов не наблюдалось (Рис. 3C).
Окрашивание серебром выделило малые зрелые нейроны и олигодендроциты. Существенных гистологических изменений в нейронах зафиксировано не было. Образование монетных столбиков из эритроцитов в периферической крови (густая кровь) было замечено в коре головного мозга спустя час после остановки сердца (рис. 3D).
При имуногистохимическом окрашивании на каспазу -3 свидетельствует о прогрессивном увеличении положительно окрашенных клеток, причем наибольшее число положительно окрашенных аксонов было зафиксировано между 6 и 9 часами. Лишь несколько активных аксонов было распознано в период между 3 и 6 часами после остановки сердца (Рис. 4A и 4B). Начиная с 9 часов после остановки сердца было зафиксировано значительное увеличение числа сильно окрашенных клеток (2.5% от всего числа нейронов коры) (Рис. 4C и 4D). Множество разрозненных положительных клеток также было отчетливо видно после 24 часов. Спустя 1 час наблюдалось неспецифичное светло-коричневое фоновое ядерное окрашивание, значение этого наблюдения остается неясным.
Рис.4

- Рис. 4A. Спустя 3 часа после остановки сердца при комнатной температуре отсутствует активизация каспазы-3 в цитоплазме нейронов коры головного мозга (иммунологическое окрашивание на каспазу-3, ×400).
- Рис. 4В. Активизация каспазы-3 в нейронах коры головного мозга (черные стрелки) спустя 3 часа после остановки сердца при комнатной температуре (иммунологическое окрашивание на каспазу-3, ×400).
- Рис. 4С. Активизация каспазы-3 в нейронах коры головного мозга (черные стрелки) спустя 9 часа после остановки сердца при комнатной температуре (иммунологическое окрашивание на каспазу-3, ×400).
- Рис. 4D. Активизация каспазы-3 в нейронах коры головного мозга (черные стрелки) спустя 12 часов после остановки сердца при комнатной температуре (иммунологическое окрашивание на каспазу-3, ×400).
Обсуждение
Электронная микроскопия аноксических повреждений мозга
В 1973 году Арсенио-Нунес и коллеги обнаружили, что после получасовой ишемии мозга, ультрамикроскопическая структура нейронов коры мозга кошек полностью восстановилась [2]. В 1974 году Хоссманн и Циммерманн спустя час после аноксии обнаружили в нейронах коры головного мозга обезьян только цитоплазматическую гомогенизацию, которая была обратима после возобновления циркуляции крови в мозге [3]. В 1975 году Клайюс и коллеги показали, что после часовой полной ишемии мозга обезьян, ультраструктура нейронов коры полностью восстанавливается [4]. В 2003 году Недзвед и коллеги также показали, что в течении 4 часов полной аноксии мозга, ультраструктура нейронов коры головного мозга человека также остается неповрежденной [11]. В своем исследовании авторы инкубировали небольшие фрагменты передней доли мозга человека, срезанные во время операции по удалению обонятельной менингиомы и изученные с помощью электронной микроскопии. Наши результаты также показали, что аутолиз нейронов мозга (клетки Пуркинье, пирамидальные нейроны гиппокампа) не приводит ни к каким ультраструктурным изменениям, так как набухание митохондрий, как это и было впервые описано 80 лет назад, остается самым общим ультраструктурным изменением после ишемии мозга [12]. Соленски и коллеги (2002) показали, что в отличие от постоянной ишемии, возобновление кровообращения стремительно приводит к серьезным тяжелым и вероятно необратимым повреждениям, в том числе к тому, что в первоначально сжатых электронно-плотных митохондриях повышается электронная плотность внутри матрикса и к аутофагии [13]. К 24 часу реперфузии митохондрии деградируют и полностью теряют форму внешней мембраны. Нейроны, находящиеся под воздействием постоянной увеличивающейся по продолжительности ишемии, также имели значительные митохондриальные повреждения, первоначально заключавшиеся в повреждениях гребешков, внутригребешковых растяжениях и потере плотности матрикса. К 24 часу постоянной ишемии, в отличии от реперфузии нейронов к тому же часу, в перинуклеарных митохондриях однородно повышена плотность матрикса, а сами митохондрии сохранили типичную округлую или трубчатую форму. Мы также наблюдали сохранение типичной округлой формы митохондрий спустя 6 часов после полной аноксии мозга при комнатной температуре.
Смерть мозга и аутолиз
В 1993 Юджихира и коллеги провели клиническое невропатологическое исследование 60 случаев смерти мозга (29 пациентов скончалось вследствие цереброваскулярного заболевания) [14]. Средняя продолжительность смерти мозга составила 99 часов. Гистология показала, что цитоплазма нейронов была бледной и еле заметной. При окрашивании миелиновые оболочки в белом веществе были бледными, а ядра глиальных клеток оказались сморщенными и пикнотическими. Во всех случаях был очевиден аутолиз мозжечкового зернистого слоя и питуитарной железы. Не наблюдалось признаков реактивного астроцитоза или инфильтрации клетками отмирающей ткани или рядом расположенных участков. Была замечена связь между степенью аутолиза и длительностью смерти мозга, но между степенью аутолиза и видом заболевания, ставшего его причиной, нет никакой связи. Наиболее заметным являлся аутолиз коры мозга, таламуса, покрышки ствола мозга, мозжечкового зернистого слоя и питуитарной железы. Клетки Пуркинье, пирамидальные клетки субполя CA1 в районе гиппокампа и пирамидальные клетки в третьем и пятом слоях коры чрезвычайно чувствительны к воздействию аноксии мозга.
В 1986 году Огата и коллеги провели анализ аутолиза зернистого слоя мозжечковой коры 45 пациентов, скончавшихся от острых цереброваскулярных заболеваний [15]. Контрольной группой были двенадцать пациентов, скончавшихся не вследствие внутричерепного заболевания. У всех пациентов, причиной смерти которых были цереброваскулярные заболевания, была обнаружена грыжа миндалин мозжечка. В центральных отделах долей мозжечка, смежных с центральным мозговым телом мозжечка, зернистый слой был более выпуклым, чем в периферийных отделах долей мозжечка. В соответствии с мнением автора, обширный аутолиз зернистого слоя, достигающий периферийных отделов долей мозжечка, может являться обнаруженной патологической характеристикой смерти мозга в том случае, когда внутричерепное кровообращение может отсутствовать или были значительно сокращено. Автор заключает, что для развития аутолиза зернистого слоя требуется меньше 1 дня. В нашем исследовании можно было увидеть первые признаки аутолитических повреждений нейронов спустя 12 часов после аноксии мозга при комнатной температуре при помощи электронной микроскопии, в то время как в срезах окрашенных гематоксилин-эозином и тионином в течении 24 часов не наблюдается аутолитических изменений. Наши результаты свидетельствуют об отсутствии аутолитических повреждений одних и тех же участков при окрашивании гематоксили-эозином и по методу Ниссля. Полученные данные дают основания предполагать, что существует взаимосвязь между наличием сосудистых заболеваний и развитием аутолитической гистопатологии.
Смерть мозга и ишемическо-реперфузиционное повреждение мозга
Почему практически невозможно реанимировать пациента после продолжительной остановки сердца при комнатной температуре? Существует два возможных объяснения. Первое - феномен "невосстановленного кровотока". Феномен "невосстановленного кровотока" был описан Амесом в 1968 году [16]. Ишемия мозга вызывает отек, который препятствует микроциркуляции из-за наружного давления на стенки капилляров. При реперфузии более крупные сосуды снова заполняются кровью, а микроциркуляция не восстанавливается из-за непроходящего отека.
Вторая причина - повреждения, вызванные свободными радикалами. Хотя ранняя реперфузия ишемической ткани важна для сохранения жизнеспособности ткани, в целом, признано, что возобновление тока крови приводит к тому, что обратимые повреждения клеток, становятся необратимыми. Свободные радикалы кислорода, образующиеся в первые моменты реперфузии, становились причиной "ишемическо-реперфузиционного повреждения" В последнее время проводилось большое количество исследований роли свободных радикалов кислорода, перекисного окисления липидов, карбоксилирование липидов, окисление белков, повреждение ДНК и повреждения,вызванные нейтрофилами, в ишемическо-реперфузиционном повреждении [17].
Смерть мозга и апоптоз
Апоптоз, или процесс запрограммированной клеточной гибели, играет важнейшую роль как во многих биологических процессах, таких как морфогенез и негативная селекция иммунной системы, так и в развитии многочисленных заболеваний, таких как рак и нейродегенеративное заболевание. Апоптоз сопровождается митохондриальной дисфункцией с последовательным выходом цитохрома с из митоходрий, в результате чего активизируется каспаза -3. Каспаза-3 играет главную роль в Fas-опосредованном апоптозе и является самой изучаемой каспазой [18]. Многочисленные исследования подтверждают, что каспаза-3 - ключевой компонент в процессе отмирания клетки, так как она блокирует кальциевые насосы клеточных мембран как нейронных, так и других соматических апоптических клеток. Блокировка кальциевого насоса каспазой ведет к значительному ионному дисбалансу, избытку кальция и, в конечном итоге, смерти клетки [19].
В 1999 г. Колбурн и коллеги показали, что не поддающееся лечению обширное ишемическое повреждение имеет некротическое, а не апоптическое строения. Однако, вероятность запрограммированных биохимических событий, ведущих к апоптозу до некроза нейронов, не может быть полностью исключена [20]. Электронная микроскопия ишемически поврежденных нейронов с или без последующей гипотермией свидетельствует о некротической, а не апоптической смерти даже тех клеток, которые погибли спустя два месяца после ишемии. Некрозу предшествовали расширение органелл и формирование внутриклеточных вакуолей. В 2002 году Шваб и коллеги показали, что степень клеточного некроза, возникшего вследствие ишемии мозга, можно снизить терапевтическим использованием ингибиторов каспазы [19]. Наши результаты показали, что в течении 9 часов после остановки сердца при комнатной температуре без реперфузии не наблюдалось очевидных признаков смерти значимых клеток. Наши выводы позволяют сделать предположение, что проведение терапии ингибиторами каспазы уменьшает степень повреждения нейронов после остановки сердца при комнатной температуре.
Механизм клеточной смерти, индуцированный истощением АТФ глубоко исследован многими учеными [21, 22]. Истощение АТФ может привести к некрозу или апоптозу [22]. В 2003 году Оно и коллеги показали, что вызванный истощением АТФ переход от апоптоза к некрозу был связан с увеличением общего объема внутриклеточных отсеков с кислотой в клетках [23]. Это увеличение, влияющее на размер лизосом, может быть причиной их повреждения. Увеличение лизосом при клеточной смерти изменяет натяжение мембран лизосом и, таким образом, повышает вероятность разрыва лизосом. В 2001 году Хишита и коллеги обнаружили, что дисфункция лизосом, вызванная истощением АТФ, приводит к высвобождению ферментов лизосом в цитозоль, и что ферменты лизосом могут быть причиной активизации каспазы-3 при апоптозе [24]. В данном исследовании, мы наблюдали первичную активизацию каспазы-3 в нейронах мозжечка и неокортекса в течении 6 часов аноксии мозга при комнатной температуре, хотя значительное увеличение количества положительно окрашенных клеток появилось лишь спустя 9 часов. Полученные данные могут быть объяснены тем, что истощение АТФ запускает программу клеточной смерти, после чего возможно высвобождение ферментов лизосом, являющееся результатом дисфункции лизосом, вызванной истощением АТФ.
Смерть мозга и гипотермия
Существует множество интересных случаев успешной реанимации людей после длительной остановки сердца (30-40 минут; утопание в холодной воде) [25, 26, 27, 28, 29, 30, 31]. Например, срезы мозга земляной белки не погибают при температуре +4°C в течении 7 дней в буферном растворе без непрерывной оксигенации [32]. Мы не проводили наш эксперимент в условиях гипотермии, так как мы были заинтересованы в изучении структурных гипоксических/ аутолитических изменений при комнатной температуре. Наш интерес к проблеме был продиктован тем фактом, что большое количество случаев остановки сердца у людей происходили при комнатной температуре или в условиях нормотермии.
В заключение, наши данные показали отсутствие серьёзных апоптических повреждений нейронов ЦНС в первые 6 часов после остановки сердца при комнатной температуре. Полученные данные могут быть полезны для судебно-медицинских экспертиз и для усовершенствования мер интенсивной терапии и реанимации. Понимание механизмов смерти мозга помогут разработать новые технологии нейро-реанимации в будущем.
Благодарности
Мы благодарим доктора Роберта К. Свитцера III, доктора философии (Ассоциация неврологии, Ноксвил, Теннеси) за проведение окрашиваний гематоксилином и эозином, по методу Ниссля, серебром и иммуногистохимических процедуры; доктора Роберта Роберсона (факультет биологии, университет штата Аризоны) за обеспечение электронной микроскопии; мистера Чарльза Дж. Казилека (группа биологической визуализации, лаборатория биовизуализация У. М. Керка, факультет биологии университет штата Аризона, Темпе, Аризона) за предоставление световой микроскопии и фотографии; мистера Билла Войса (фонд продления жизни Alcor, Скоттсдейл, Аризона) за оказанную помощь при проведении экспериментов, доктора Уильяма Д. Андерсона (отделение патологий, госпиталь и медицинский центр св. Иосифа, Феникс, Аризона) за консультации. Данный исследовательский проект был проведен при поддержке фонда продления жизни Alcor (Скоттсдейл, Аризона 85260).
Пожалуйста, всю корреспонденцию отправляйте доктору Сергею В. Шелегу, фонд продления жизни Alcor, Скоттсдейл, Аризона 85260, США.
Ссылки:
1. Walker EA. Cerebral Death. 3rd Ed. Baltimore, MD and Urban and Schwartzenberg GmbH: 1985.
2. Arsenio-Nunes ML, Hossmann KA, Farkas-Bargeton E. Ultrastructural and histochemical investigation of the cerebral cortex of cat during and after complete ischaemia. Acta Neuropathol (Berl) 1973;26:329–344. [PubMed]
3. Hossmann KA, Zimmermann V. Resuscitation of the monkey brain after 1h complete ischemia. I. Physiological and morphological observations. Brain Res. 1974;29:59–74. [PubMed]
4. Kleihues P, Hossmann KA, Pegg AE, Kobayashi K, Zimmermann V. Resuscitation of the monkey brain after one hour complete ischemia. III. Indications of metabolic recovery. Brain Res. 1975;95:61–73.[PubMed]
5. Zimmermann V, Hossmann KA. Resuscitation of the monkey brain after one hour's complete ischemia. II. Brain water and electrolytes. Brain Res. 1975;85:1–11. [PubMed]
6. Rothstein RP, Levison SW. Gray matter oligodendrocyte progenitors and neurons die caspase-3 mediated deaths subsequent to mild perinatal hypoxic/ischemic insults. Dev Neurosci. 2005;27:149–159.[PMC free article] [PubMed]
7. Iwata O, Iwata S, Tamura M, Nakamura T, Sugiura M, Ogiso Y, Takashima S. Early head cooling in newborn piglets is neuroprotective even in the absence of profound systemic hypothermia. Pediatr Int.2003;45:522–529. [PubMed]
8. Zhu H, Meloni BP, Bojarski C, Knuckey MW, Knuckey NW. Post-ischemic modest hypothermia (35°C) combined with intravenous magnesium is more effective at reducing CA1 neuronal death than either treatment used alone following global cerebral ischemia in rats. Exp Neurol. 2005;193:361–368. [PubMed]
9. Vereczki V, Martin E, Rosenthal RE, Hof PR, Hoffman GE, Fiskum G. Normoxic resuscitation after cardiac arrest protects against hippocampal oxidative stress, metabolic dysfunction, and neuronal death. J Cereb Blood Flow Metab. 2006;26:821–835. [PMC free article] [PubMed]
10. de Olmos JS, Beltramino CA, de Olmos de Lorenzo S. Use of an amino-cupric-silver technique for the detection of early and semiacute neuronal degeneration caused by neurotoxicants, hypoxia, and physical trauma. Neurotoxicol Teratol. 1994;16:545–561. [PubMed]
11. Nedzved MK, Sheleg SV, Gerasimova TN, Oleshkevich FV, Wolf N. [Morphologic changes of human cerebral cortex neurons under long duration anoxia] Zdravoohranenie. 2003;7:10–12. (in Russian)
12. Spielmeyer W. Histopathologie des Nervensystems. Berlin, Germany: Springer; 1922. pp. 74–79.
13. Solenski NJ, diPierro CG, Trimmer PA, Kwan AL, Helm GA. Ultrastructural changes of neuronal mitochondria after transient and permanent cerebral ischemia. Stroke. 2002;33:816–824. [PubMed]
14. Ujihira N, Hashizume Y, Takahashi A. A clinico-neuropathological study on brain death. Nagoya J Med Sci. 1993;56:89–99. [PubMed]
15. Ogata J, Yutani C, Imakita M, Ueda H, Waki R, Ogawa M, Yamaguchi T, Sawada T, Kikuchi H. Autolysis of the granular layer of the cerebellar cortex in brain death. Acta Neuropathol (Berl) 1986;70:75–78. [PubMed]
16. Ames A, 3rd, Wright RL, Kowada M, Thurston JM, Majno G. Cerebral ischemia. II. The no-reflow phenomenon. Am J Pathol. 1968;52:437–453. [PMC free article] [PubMed]
17. іşlekel H, іşlekel S, Güner G. Biochemical mechanism and tissue injury of cerebral ischemia and reperfusion. Part II: Tissue injury. NOROL BIL D 2000, 17 (Electronic publication). Available at:http://www.med.ege.edu.tr/norolbil/2000/NBD09200.html. Accessed January 06, 2006.
18. Woo M, Hakem A, Elia AJ, Hakem R, Duncan GS, Patterson BJ, Mak TW. In vivo evidence that caspase-3 is required for Fas-mediated apoptosis of hepatocytes. J Immunol. 1999;163:4909–4916.[PubMed]
19. Schwab BL, Guerini D, Didszun C, Bano D, Ferrando-May E, Fava E, Tam J, Xu D, Xanthoudakis S, Nicholson DW, Carafoli E, Nicotera P. Cleavage of plasma membrane calcium pumps by caspases: a link between apoptosis and necrosis. Cell Death Differ. 2002;9:818–831. [PubMed]
20. Colbourne F, Sutherland GR, Auer RN. Electron microscopic evidence against apoptosis as the mechanism of neuronal death in global ischemia. J Neurosci. 1999;19:4200–4210. [PubMed]
21. Kelly KJ, Plotkin Z, Dagher PC. Guanosine supplementation reduces apoptosis and protects renal function in the setting of ischemic injury. J Clin Invest. 2001;108:1291–1298. [PMC free article] [PubMed]
22. Lieberthal W, Menza SA, Levine JS. Graded ATP depletion can cause necrosis or apoptosis of cultured mouse proximal tubular cells. Am J Physiol. 1998;274:F315–327. [PubMed]
23. Ono K, Kim SO, Han J. Susceptibility of lysosomes to rupture is a determinant for plasma membrane disruption in tumor necrosis factor alpha-induced cell death. Mol Cell Biol. 2003;23:665–676.[PMC free article] [PubMed]
24. Hishita T, Tada-Oikawa S, Tohyama K, Miura Y, Nishihara T, Tohyama Y, Yoshida Y, Uchiyama T, Kawanishi S. Caspase-3 activation by lysosomal enzymes in cytochrome c-independent apoptosis in myelodysplastic syndrome-derived cell line P39. Cancer Res. 2001;61:2878–2884. [PubMed]
25. Chochinov AH, Baydock BM, Bristow GK, Giesbrecht GG. Recovery of a 62-year-old man from prolonged cold water submersion. Ann Emerg Med. 1998;31:127–131. [PubMed]
26. Felix WR, Jr, MacDonnel KF, Jacobs L. Resuscitation from drowning in cold water. N Engl J Med.1981;304:843–844. [PubMed]
27. Graf D, Meier P, Guse HG, Leitz KH, Bachmann H. [A drowning accident of long duration with deep hypothermia and rewarming with extracorporeal circulation. A report of 2 patients] Monatsschr Kinderheilkd. 1989;137:415–418. (in German) [PubMed]
28. Jacobsen JB, Nielsen H, Andersen PK. Resuscitation from drowning in cold water. N Engl J Med.1981;305:580–581. [PubMed]
29. Kugler-Podelleck I, Rodewald G, Horatz K, Kugler S, Muller-Brunotte P. Resuscitation after drowning in freezing water. Ger Med Mon. 1966;11:232–236. [PubMed]
30. Modell JH, Idris AH, Pineda JA, Silverstein JH. Survival after prolonged submersion in freshwater in Florida. Chest. 2004;125:1948–1951. [PubMed]
31. Norberg WJ, Agnew RF, Brunsvold R, Sivanna P, Browdie DA, Fisher D. Successful resuscitation of a cold water submersion victim with the use of cardiopulmonary bypass. Crit Care Med. 1992;20:1355–1357. [PubMed]
32. Pakhotin PI, Belousov AB, Otmakhov NA. [A comparative analysis of the functional stability (based on electrophysiological criteria) of slices of the suslik and guinea pig brains maintained under deep hypothermia] Zh Evol Biokhim Fiziol. 1991;27:479–485. (in Russian) [PubMed]
-------------------------------------------------------------------------------
http://translatedby.com/you/stability-and-autolysis-of-cortical-neurons-...
Original (English): Stability and Autolysis of Cortical Neurons in Post-Mortem Adult Rat Brains
Translation: © Анечка, Blockaderunner.











































